

Methane production and rumen-protected fats


Methane has a much shorter life-time than CO2 in the atmosphere, which makes it an attractive target for shorter-term gains in global warming reduction. Rumen-protected fat supplements offer a largely unique approach to methane mitigation efforts.
Methane (CH4) is one of the 3 main greenhouse gases along with carbon dioxide (CO2) and nitrous oxide (N2O). It is highly potent, having a global warming potential 28 times that of CO2. Total global greenhouse gas emissions from livestock are estimated to account for 14.5% of total anthropogenic emissions and 40% of this is accounted for by enteric methane from ruminants (from Beauchemin et al., 2020). Most (approx. 90%) of the enteric methane has its origin in the rumen through the process of microbial methanogenesis (Martin et al., 2010).
However, methane has a much shorter life-time than CO2 in the atmosphere (half-life; 8.6 years; Muller and Muller, 2017), which makes it an attractive target for shorter-term gains in global warming reduction (Beauchemin et al., 2020).
Energy loss
In addition to environmental implications, ruminal methane production also represents a considerable loss of energy to the animal, ranging from 2-12% of gross energy intake of ruminants (Johnson and Johnson, 1995), though values between 3% and 7% are more realistic in intensive dairy production (Martin et al., 2008).
Methane production by ruminants
Methane is produced as a by-product of fermentation of feed components by rumen microbiota (bacteria, protozoa and fungi) to volatile fatty acids (VFA) in the rumen. It is strongly influenced by dry matter intake, which determines volume of fermentable feed substrates, and fermentability of feed.
The fermentation process involves oxidation of reduced co-factors NADH, NADPH and FADH through dehydrogenation reactions resulting in release of hydrogen (H), as well as CO2 (Martin et al., 2010). This hydrogen is then used by methanogenic archaea to reduce CO2 via the hydrogenotrophic pathway (Beauchemin et al., 2020) leading to formation of methane, as follows :
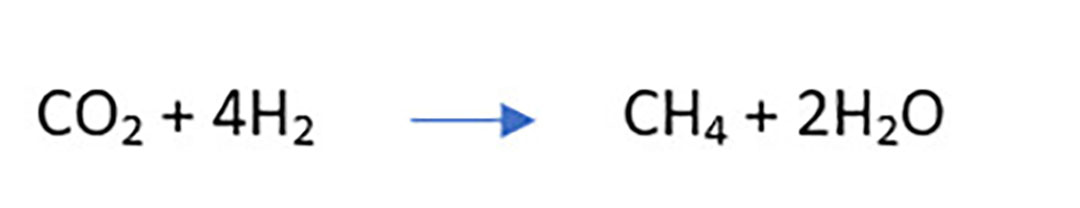
Methane gas produced is eructated (burped out) by the animal, facilitating removal of excess hydrogen from the rumen. This is a natural process essential to the continuation of anaerobic rumen fermentation and microbial growth (from van Zijderveld et al., 2011).
Effect of fermentative energy source in the rumen
Fermentation of different energy sources in the rumen leads to alternative patterns of VFA production. Fermentation of fibrous material in the rumen drives acetate and butyrate which lead to a net release of hydrogen, whereas increasing proportion of concentrates in the ration and development of starch-fermenting microbes leads to increased propionate production (an alternative hydrogen sink) and reduced hydrogen release for methane production (Martin et al., 2010). Furthermore, increasing concentrate supplementation and starch intake can decrease ruminal pH leading to inhibition of methanogenic growth (Beauchemin et al., 2020).
Dietary fat
In contrast to other nutrient sources, dietary fat is unique in that it is not fermented to VFA in the rumen. This characteristic offers a specific method of increasing energy supply without adding to fermentation-induced hydrogen production in the rumen.
Beauchemin et al. (2020) summarised data from meta-analyses and reported a reduction in methane production (g/d) of between 1% and 5% per 1% increase in dietary fat concentration. Mechanisms include replacement of fermentable organic matter, inhibition of protozoa and methanogens, acting as a hydrogen sink through biohydrogenation, and reduction of fibre digestibility.
However, the potential for methane mitigation using ‘rumen-active’ lipids is limited due to negative effects on animal production. In particular, the risks of reduced fibre digestibility, feed efficiency and formation of trans fatty acid isomers that cause milk fat depression with increasing rumen-active fat sources need to be considered (Palmquist and Jenkins, 2017).
Rumen-protected fat supplements
In contrast, rumen-protected fats were developed to overcome the negative effects on rumen fermentation and fibre digestibility associated with unsaturated ‘rumen-active’ oils (Palmquist and Jenkins, 2017). Their development enabled supplementation of ruminant diets with lipid to increase energy density without the negative intra-ruminal and production effects associated with ‘free’ oils, with additional benefits including the ability to deliver a greater proportion of biologically-active unsaturated fatty acids (e.g. C18:1, omega-3) through the rumen to the small intestine.
Rumen-protected fat supplements offer a largely unique approach to methane mitigation efforts. As established ingredients for improving production and fertility aspects in dairy and other ruminant diets, a beneficial effect on methane reduction is achieved more as a ‘side-effect’ in addition to the performance gains for which these supplements are typically used for economic return. With minimal effect on ruminal environment or microbiota, the primary mechanism for rumen-protected fat-mediated reduction of methane is via direct replacement of fermentable organic matter in the diet to reduce substrate availability for hydrogen production.
Established research work from the US evaluated the effects of replacing 2.95% ground maize with rumen-protected fat (Megalac; calcium salts of fatty acids) (targeted to supply approx. 454 g of fat supplement/lactating cow/day) (Table 1). Fat supplementation increased milk yield by 2.3 kg/cow/d and reduced daily methane production by 7.5%, translating to a 13.7% reduction in methane intensity. Non-lactating cows had a tendency for lower methane per unit of gross energy intake (-5.1%).
Persistency of effects
Effect of methane inhibitors can be influenced by the risk of adaptation of ruminal microflora over time. However, this is unlikely to occur where methane is reduced by the addition of rumen-protected fat to a ration given that the primary mechanism involves a reduction in fermentable organic matter in the rumen, and effects are hypothesised to be persistent (Alstrup et al., 2015).
Fertility effects
Rumen-protected fats with specific fatty acid profiles are also commonly targeted to improve fertility, with mechanisms including increasing energy supply, increasing progesterone production, and improving quality and survivability of fertilised eggs. Improved fertility traits can translate to fewer replacement animals needed, which has major implications on methane production at farm level. Garnsworthy (2011) determined that the proportion of total methane emissions produced by dairy replacements can be reduced from 30% at poor fertility levels (oestrous detection 50%; conception rate 30%) to 10% under improved fertility conditions (oestrous detection 70%; conception rate 60%).
Summary
Rumen-protected fats are widely used in dairy and other ruminant diets to provide an economic return through enhanced productivity effects. The indirect effect on methane reduction should be considered as an additional benefit without additional costs to the producer, and with expected persistent effects through the period of use.
Join 13,000+ subscribers
Subscribe to our newsletter to stay updated about all the need-to-know content in the dairy sector, two times a week.







